訓練されていない目には、植物は衝動的に成長しているように見え、葉をランダムに飛び出して大きな緑色のごちゃ混ぜにします。 しかし、よく見てみると、竹の実のバランスの取れた対称性から多肉植物の魅惑的なスパイラルまで、自然界のいたるところに奇妙な規則的なパターンがいくつか現れていることがわかります。
実際、これらのパターンは十分に一貫しているため、寒くて難しい数学は有機的成長をかなりよく予測できます。 葉序、または葉のパターンの研究の中心となっている前提の1つは、葉が自分の空間を保護するというものです。 すでに存在する葉は新しい葉に抑制的な影響を与え、他の人が近くで成長するのを防ぐ信号を発するという考えに基づいて、科学者は自然の多くの一般的なデザインをうまく再現できるモデルを作成しました。 たとえば、魅力的なフィボナッチシーケンスは、ヒマワリの種の配置からオウムガイの殻、松ぼっくりまで、あらゆるものに現れます。 現在のコンセンサスでは、成長ホルモンオーキシンと植物全体にそれを輸送するタンパク質の動きがそのようなパターンの原因であるということです。
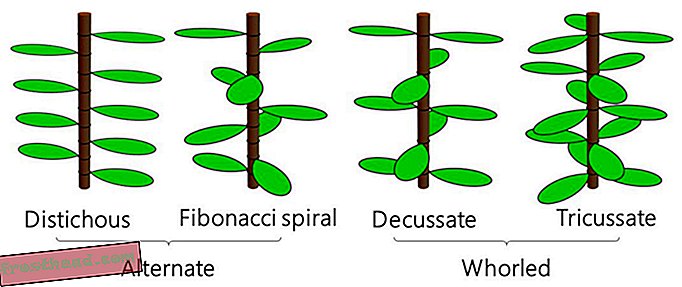 ノードごとに1つの葉を持つ葉の配置は、交互葉序と呼ばれ、ノードごとに2つ以上の葉を持つ配置は、渦巻き葉序と呼ばれます。 一般的な代替タイプは、フィステルフィス(竹)とフィボナッチスパイラルフィロタクシス(多肉植物のスパイラルアロエ)であり、一般的な渦巻きタイプは、葉状のフィロタクシス(バジルまたはミント)と三日月状のフィロタキシー(ネメリウムオレアンダー 、時々dogbaneとも呼ばれます)です。 (CC-BY-NDの米倉貴明)
ノードごとに1つの葉を持つ葉の配置は、交互葉序と呼ばれ、ノードごとに2つ以上の葉を持つ配置は、渦巻き葉序と呼ばれます。 一般的な代替タイプは、フィステルフィス(竹)とフィボナッチスパイラルフィロタクシス(多肉植物のスパイラルアロエ)であり、一般的な渦巻きタイプは、葉状のフィロタクシス(バジルまたはミント)と三日月状のフィロタキシー(ネメリウムオレアンダー 、時々dogbaneとも呼ばれます)です。 (CC-BY-NDの米倉貴明) ただし、1990年代以降支配的なDouady方程式とCouder方程式(DC1およびDC2として知られている)を含む、特定のリーフアレンジメントは、植物成長の一般的なモデルを妨害し続けています。 Orixa japonicaとして知られる低木を研究している東京大学の研究者が率いるチームは、以前の方程式では植物の異常な構造を再現できなかったため、モデル自体を再考することにしました。 PLOS Computational Biologyの新しい研究で説明されている彼らの更新されたモデルは、かつては解明されなかったパターンを再現するだけでなく、以前の方程式よりも他のより一般的な配置をよりよく記述するかもしれない、と著者は言います。
「ほとんどの植物では、葉序パターンは対称性(らせん対称性または放射状対称性)を持っています」と、東京大学の植物生理学者杉山宗孝(新しい研究の主著者)は言います。 「しかし、この特別な植物Orixa japonicaでは、葉序パターンは対称ではなく、非常に興味深いものです。 10年以上前に、各葉原基の抑制力の変化がこの特異なパターンを説明するかもしれないという考えに至りました。」
植物学者は、発散角、または連続した葉の間の角度を使用して、植物の葉序を定義します。 ほとんどのリーフアレンジメントパターンは一定の発散角を維持しますが、日本および東アジアの他の地域に自生するO. japonica低木は、180度、90度、180度の4つの繰り返し角度の交互のシリーズで葉を成長させます。その後270度。
 葉のさまざまな広がり角が見えるオリックス・ジャポニカの低木。 (CC BY-SA 4.0でのWikicommons経由のQwert1234)
葉のさまざまな広がり角が見えるオリックス・ジャポニカの低木。 (CC BY-SA 4.0でのWikicommons経由のQwert1234) 研究者が「オリックス酸塩」の葉序と呼んだこのパターンは、他の分類群の植物(「赤熱ポーカー」花のKniphofia uvariaやクレープギンバイカLagerstroemia indicaなど )が同じ葉を交互に配置するため、一度限りの異常ではありません複雑なシーケンス。 葉の配置は進化ツリーのさまざまな場所に現れるため、著者は類似性はさらなる研究を必要とする共通のメカニズムに由来すると結論付けました。
異なるパラメーターでDouadyとCouderの方程式をテストした後、著者は交互のオリックスの配置に近いパターンを作成できましたが、シミュレートされた植物はいずれも、解剖し研究したO. japonicaサンプルと完全に一致しませんでした。 そこで、チームはDouadyおよびCouderの方程式に別の変数を追加することで新しいモデルを構築しました。葉の年齢です。 以前のモデルでは、葉の抑制力は時間とともに変わらないと想定されていましたが、この定数は「生物学の観点からは自然ではありませんでした」と杉山は言います。 代わりに、杉山のチームは、これらの「キープアウェイ」信号の強度が時間とともに変化する可能性を考慮しました。
結果として得られたモデル(チームは拡張DouadyモデルとCouderモデル、EDC1およびEDC2と呼びます)は、コンピューター化された成長を通じて、O。japonicaの複雑な葉の配置を再現することに成功しました。 この偉業を超えて、拡張された方程式は、他のすべての一般的な葉のパターンも生成し、これらの品種の固有振動数を以前のモデルよりも正確に予測しました。 特にスパイラルパターンの植物の場合、新しいEDC2モデルは他のアレンジメントと比較してフィボナッチスパイラルの「スーパー優位性」を予測しましたが、以前のモデルはこの特定の形状が自然の至る所に現れる理由を説明できませんでした。
「当社のモデル、EDC2は、すべての主要なタイプの葉序に加えてオリックスパターンを生成できます。 これは明らかに以前のモデルよりも優れています」と杉山氏は言います。 「EDC2は、さまざまなパターンの自然発生にも適しています。」
 オリックスジャポニカの枝の葉(左上)とオリックスの葉序の模式図(右)。 orixateパターンは、葉の間の角度の特異な4サイクルの変化を示します。 走査型電子顕微鏡の画像(中央と左下)は、葉が最初に成長し始めるO. japonicaの冬芽を示しています。 原始葉には、P8として最も古い葉、P1として最も若い葉で順番にラベルが付けられます。 ラベルOは、シュートの頂点を示します。 (米倉貴明/岩本明俊/杉山宗孝監督下)
オリックスジャポニカの枝の葉(左上)とオリックスの葉序の模式図(右)。 orixateパターンは、葉の間の角度の特異な4サイクルの変化を示します。 走査型電子顕微鏡の画像(中央と左下)は、葉が最初に成長し始めるO. japonicaの冬芽を示しています。 原始葉には、P8として最も古い葉、P1として最も若い葉で順番にラベルが付けられます。 ラベルOは、シュートの頂点を示します。 (米倉貴明/岩本明俊/杉山宗孝監督下) 著者らは、葉の年齢がこれらの成長パターンに正確に影響する原因をまだ結論付けることはできませんが、杉山は植物の成長過程でオーキシン輸送システムの変化に関係していると推測しています。
このような謎は、計算モデルと実験室実験の間の「プッシュアンドプル」によって解決できると、研究に関与していない計算生物学者であるCiera Martinez氏は言います。 著者のモデルは、葉序のより良い理解への刺激的なステップを提供し、他の植物学者が植物の解剖と分析でギャップを埋める余地を残します。
「モデルの場合、正確なメカニズムはまだわからないかもしれませんが、少なくとも何を探すべきかについての強力な手がかりが与えられています」とMartinez氏はメールで述べています。 「今、実際の植物の分子メカニズムを詳しく調べて、モデルが予測するものを発見する必要があります。」
 新しい葉(赤い半円)が芽の頂点(中央の黒い円)から形成され、外側に向かって成長するときの「オリックス」葉序における葉の配置パターンのトップダウンビュー(CC-BY-NDの米倉貴明)
新しい葉(赤い半円)が芽の頂点(中央の黒い円)から形成され、外側に向かって成長するときの「オリックス」葉序における葉の配置パターンのトップダウンビュー(CC-BY-NDの米倉貴明) 杉山のチームは、モデルをさらに改良し、既知のすべてのフィロタクティックパターンを生成するように取り組んでいます。 杉山は葉の多いコードをクラックしそうだと考えていますが、1つの「神秘的な」葉のパターン、小さな発散角を持つらせんは、依然として計算の予測を回避します。
「私たちの研究が社会にとって実際に役立つとは思わない」と杉山は言う。 「しかし、それが自然の対称的な美しさの理解に役立つことを願っています。」